
Importance of Cry Proteins in Biotechnology: Initially a Bioinsecticide, Now a Vaccine Adjuvant

 ,
,
Abstract
:1. Introduction
2. Bacillus Thuringiensis (Bt)
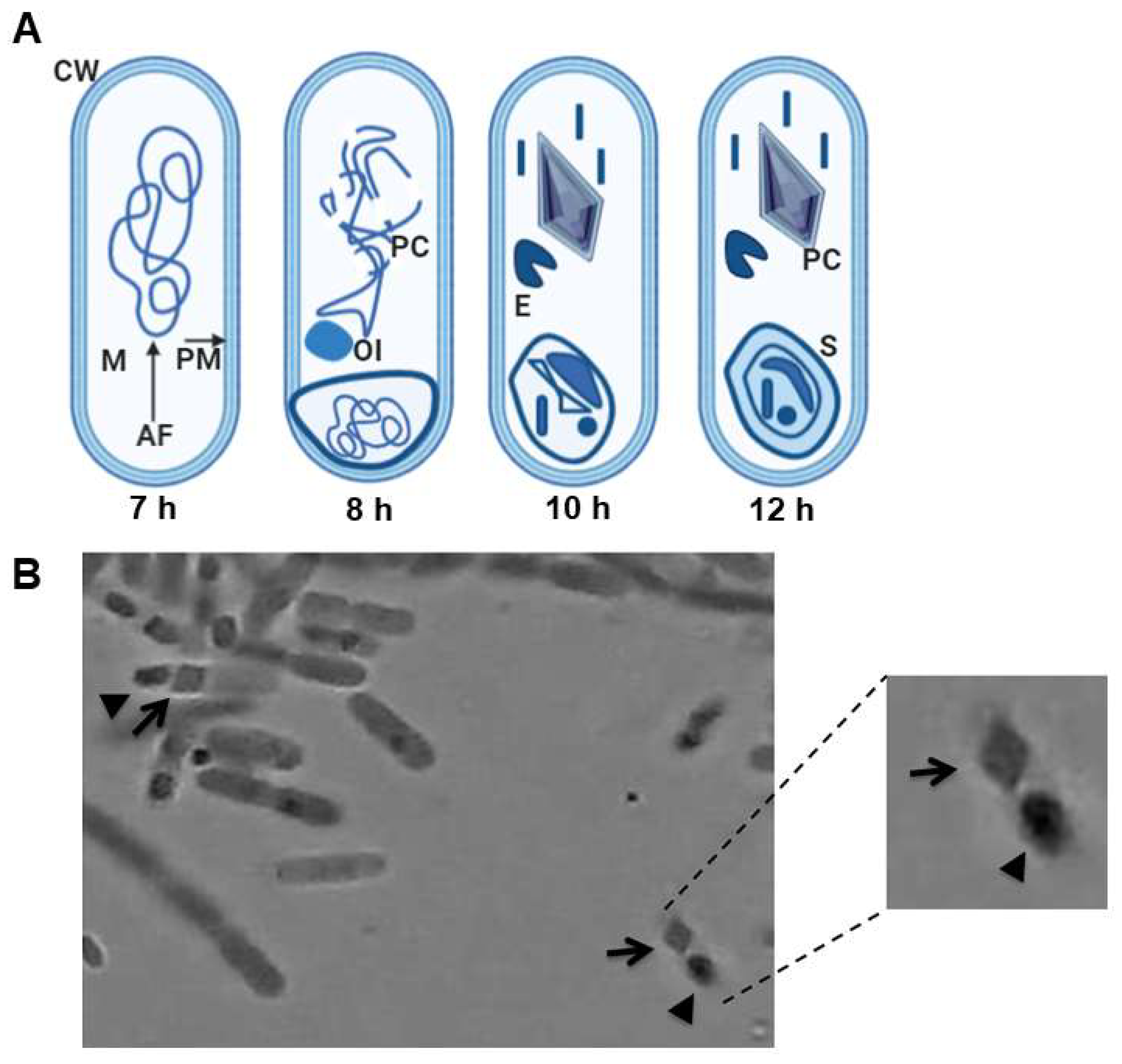
2.1. Overview of Bt and Spore Formation
2.2. Cry Proteins
2.3. Action Mechanism of Cry as a Bioinsecticide
3. Adjuvants Currently in Use
3.1. Benefits of Cry Relative to Other Adjuvants
3.2. An Overview of Cry as a Natural Adjuvant
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Apostolico Jde, S.; Lunardelli, V.A.; Coirada, F.C.; Boscardin, S.B.; Rosa, D.S. Adjuvants: Classification, Modus Operandi, and Licensing. J. Immunol. Res. 2016, 2016, 1459394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcon, N.; Di Pasquale, A. From discovery to licensure, the Adjuvant System story. Hum. Vaccin. Immunother. 2017, 13, 19–33. [Google Scholar] [CrossRef]
- Sjoblad, R.D.; McClintock, J.T.; Engler, R. Toxicological considerations for protein components of biological pesticide products. Regul. Toxicol. Pharmacol. 1992, 15, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Koch, M.S.; Ward, J.M.; Levine, S.L.; Baum, J.A.; Vicini, J.L.; Hammond, B.G. The food and environmental safety of Bt crops. Front. Plant. Sci. 2015, 6, 283. [Google Scholar] [CrossRef]
- Mezzomo, B.P.; Miranda-Vilela, A.L.; Grisolia, C.K. Toxicological Evaluation of a Potential Immunosensitizer for Use as a Mucosal Adjuvant--Bacillus thuringiensis Cry1Ac Spore-Crystals: A Possible Inverse Agonist that Deserves Further Investigation. Toxins 2015, 7, 5348–5358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.A.; Griko, N.; Junker, M.; Bulla, L.A. Bacillus thuringiensis: A genomics and proteomics perspective. Bioeng. Bugs 2010, 1, 31–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Barjac, H.; Bonnefoi, A. A classification of strains of Bacillus thuringiensis Berliner with a key to their differentiation. J. Invertebr. Pathol. 1968, 11, 335–347. [Google Scholar] [CrossRef]
- Fast, P.G.; Angus, T.A. Effects of Parasporal Inclusions of Bacillus Thuringiensis Var. Sotto Ishiwata on the Permeability of the Gut Wall of Bompyx Mori (Linnaeus) Larvae. J. Invertebr. Pathol. 1965, 20, 29–32. [Google Scholar] [CrossRef]
- Fast, P.G.; Donaghue, T.P. The delta-endotoxin of Bacillus thuringiensis. II. On the mode of action. J. Invertebr. Pathol. 1971, 18, 135–138. [Google Scholar] [CrossRef]
- Jouzani, G.S.; Valijanian, E.; Sharafi, R. Bacillus thuringiensis: A successful insecticide with new environmental features and tidings. Appl. Microbiol. Biotechnol. 2017, 101, 2691–2711. [Google Scholar] [CrossRef]
- Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.Z.; Hale, K.; Carta, L.; Platzer, E.; Wong, C.; Fang, S.C.; Aroian, R.V. Bacillus thuringiensis crystal proteins that target nematodes. Proc. Natl. Acad. Sci. USA 2003, 100, 2760–2765. [Google Scholar] [CrossRef] [Green Version]
- Lambert, B.; Hofte, H.; Annys, K.; Jansens, S.; Soetaert, P.; Peferoen, M. Novel Bacillus thuringiensis insecticidal crystal protein with a silent activity against coleopteran larvae. Appl. Environ. Microbiol. 1992, 58, 2536–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, B. The biology, ecology and taxonomy of Bacillus thuringiensis and related bacteria. In Bacillus thuringiensis and Lysinibacillus sphaericus; Springer: Berlin/Heidelberg, Germany, 2017; pp. 19–39. [Google Scholar]
- Hofte, H.; de Greve, H.; Seurinck, J.; Jansens, S.; Mahillon, J.; Ampe, C.; Vandekerckhove, J.; Vanderbruggen, H.; van Montagu, M.; Zabeau, M.; et al. Structural and functional analysis of a cloned delta endotoxin of Bacillus thuringiensis berliner 1715. Eur. J. Biochem. 1986, 161, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Key, S.; Ma, J.K.; Drake, P.M. Genetically modified plants and human health. J. R. Soc. Med. 2008, 101, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Hofte, H.; Seurinck, J.; Van Houtven, A.; Vaeck, M. Nucleotide sequence of a gene encoding an insecticidal protein of Bacillus thuringiensis var. tenebrionis toxic against Coleoptera. Nucleic. Acids. Res. 1987, 15, 7183. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, P.; Banerjee, G. Recent advancement on chemical arsenal of Bt toxin and its application in pest management system in agricultural field. 3 Biotech. 2018, 8, 201. [Google Scholar] [CrossRef]
- Christou, P.; Capell, T.; Kohli, A.; Gatehouse, J.A.; Gatehouse, A.M. Recent developments and future prospects in insect pest control in transgenic crops. Trends. Plant. Sci. 2006, 11, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Romeis, J.; Meissle, M.; Bigler, F. Transgenic crops expressing Bacillus thuringiensis toxins and biological control. Nat. Biotechnol. 2006, 24, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Valtierra-de-Luis, D.; Villanueva, M.; Berry, C.; Caballero, P. Potential for Bacillus thuringiensis and Other Bacterial Toxins as Biological Control Agents to Combat Dipteran Pests of Medical and Agronomic Importance. Toxins 2020, 12, 773. [Google Scholar] [CrossRef]
- Bernhard, K. Quantitative determination of delta-endotoxin contents in spray-dried preparations ofBacillus thuringiensis strain GC-91. World J. Microbiol. Biotechnol. 1992, 8, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.A.; Travers, R.S. Worldwide Abundance and Distribution of Bacillus thuringiensis Isolates. Appl. Environ. Microbiol. 1989, 55, 2437–2442. [Google Scholar] [CrossRef] [Green Version]
- Aronson, A.I.; Beckman, W.; Dunn, P. Bacillus thuringiensis and related insect pathogens. Microbiol. Rev. 1986, 50, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Ichimatsu, T.; Mizuki, E.; Nishimura, K.; Akao, T.; Saitoh, H.; Higuchi, K.; Ohba, M. Occurrence of Bacillus thuringiensis in fresh waters of Japan. Curr. Microbiol. 2000, 40, 217–220. [Google Scholar] [CrossRef]
- Hofte, H.; Whiteley, H.R. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 1989, 53, 242–255. [Google Scholar] [CrossRef]
- Bulla, L.A., Jr.; Bechtel, D.B.; Kramer, K.J.; Shethna, Y.I.; Aronson, A.I.; Fitz-James, P.C. Ultrastructure, physiology, and biochemistry of Bacillus thuringiensis. Crit. Rev. Microbiol. 1980, 8, 147–204. [Google Scholar] [CrossRef] [PubMed]
- Bechtel, D.B.; Bulla, L.A., Jr. Ultrastructural analysis of membrane development during Bacillus thuringiensis sporulation. J. Ultrastruct. Res. 1982, 79, 121–132. [Google Scholar] [CrossRef]
- Agaisse, H.; Lereclus, D. How does Bacillus thuringiensis produce so much insecticidal crystal protein? J. Bacteriol. 1995, 177, 6027–6032. [Google Scholar] [CrossRef] [Green Version]
- Baum, J.A.; Malvar, T. Regulation of insecticidal crystal protein production in Bacillus thuringiensis. Mol. Microbiol. 1995, 18, 1–12. [Google Scholar] [CrossRef]
- Lopez-Meza, J.E.; Ibarra, J.E. Characterization of a Novel Strain of Bacillus thuringiensis. Appl. Environ. Microbiol. 1996, 62, 1306–1310. [Google Scholar] [CrossRef] [Green Version]
- Fiuza, L.M.; Schunemann, R.; Pinto, L.M.; Zanettini, M.H. Two new Brazilian isolates of Bacillus thuringiensis toxic to Anticarsia gemmatalis (Lepidoptera: Noctuidae). Braz. J. Biol. 2012, 72, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Mukhija, B.; Khanna, V. Isolation, characterization and crystal morphology study of Bacillus thuringiensis isolates from soils of Punjab. J. Pure Appl. Microbiol. 2018, 12, 189–194. [Google Scholar] [CrossRef]
- Schnepf, H.E.; Whiteley, H.R. Cloning and expression of the Bacillus thuringiensis crystal protein gene in Escherichia coli. Proc. Natl. Acad. Sci. USA 1981, 78, 2893–2897. [Google Scholar] [CrossRef] [Green Version]
- Crickmore, N.; Berry, C.; Panneerselvam, S.; Mishra, R.; Connor, T.R.; Bonning, B.C. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2020, 107438. [Google Scholar] [CrossRef] [PubMed]
- Crickmore, N.; Zeigler, D.R.; Feitelson, J.; Schnepf, E.; Van Rie, J.; Lereclus, D.; Baum, J.; Dean, D.H. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 807–813. [Google Scholar] [CrossRef] [Green Version]
- Crickmore, N.; Baum, J.; Bravo, A.; Lereclus, D.; Narva, K.; Sampson, K.; Schnepf, E.; Sun, M.; Zeigler, D.R. Bacillus thuringiensis toxin nomenclature. Available online: http://www.btnomenclature.info/ (accessed on 18 September 2021).
- Boonserm, P.; Mo, M.; Angsuthanasombat, C.; Lescar, J. Structure of the functional form of the mosquito larvicidal Cry4Aa toxin from Bacillus thuringiensis at a 2.8-angstrom resolution. J. Bacteriol. 2006, 188, 3391–3401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.D.; Carroll, J.; Ellar, D.J. Crystal structure of insecticidal delta-endotoxin from Bacillus thuringiensis at 2.5 A resolution. Nature 1991, 353, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Grochulski, P.; Masson, L.; Borisova, S.; Pusztai-Carey, M.; Schwartz, J.L.; Brousseau, R.; Cygler, M. Bacillus thuringiensis CryIA(a) insecticidal toxin: Crystal structure and channel formation. J. Mol. Biol. 1995, 254, 447–464. [Google Scholar] [CrossRef]
- Galitsky, N.; Cody, V.; Wojtczak, A.; Ghosh, D.; Luft, J.R.; Pangborn, W.; English, L. Structure of the insecticidal bacterial delta-endotoxin Cry3Bb1 of Bacillus thuringiensis. Acta Crystallogr. D Biol. Crystallogr. 2001, 57, 1101–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonserm, P.; Davis, P.; Ellar, D.J.; Li, J. Crystal structure of the mosquito-larvicidal toxin Cry4Ba and its biological implications. J. Mol. Biol. 2005, 348, 363–382. [Google Scholar] [CrossRef]
- Guo, S.; Ye, S.; Liu, Y.; Wei, L.; Xue, J.; Wu, H.; Song, F.; Zhang, J.; Wu, X.; Huang, D.; et al. Crystal structure of Bacillus thuringiensis Cry8Ea1: An insecticidal toxin toxic to underground pests, the larvae of Holotrichia parallela. J. Struct. Biol. 2009, 168, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Hui, F.; Scheib, U.; Hu, Y.; Sommer, R.J.; Aroian, R.V.; Ghosh, P. Structure and glycolipid binding properties of the nematicidal protein Cry5B. Biochemistry 2012, 51, 9911–9921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derbyshire, D.J.; Ellar, D.J.; Li, J. Crystallization of the Bacillus thuringiensis toxin Cry1Ac and its complex with the receptor ligand N-acetyl-D-galactosamine. Acta Crystallogr. D Biol. Crystallogr. 2001, 57, 1938–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morse, R.J.; Yamamoto, T.; Stroud, R.M. Structure of Cry2Aa suggests an unexpected receptor binding epitope. Structure 2001, 9, 409–417. [Google Scholar] [CrossRef]
- Sawaya, M.R.; Cascio, D.; Gingery, M.; Rodriguez, J.; Goldschmidt, L.; Colletier, J.P.; Messerschmidt, M.M.; Boutet, S.; Koglin, J.E.; Williams, G.J.; et al. Protein crystal structure obtained at 2.9 A resolution from injecting bacterial cells into an X-ray free-electron laser beam. Proc. Natl. Acad. Sci. USA 2014, 111, 12769–12774. [Google Scholar] [CrossRef] [Green Version]
- Jing, X.; Yuan, Y.; Wu, Y.; Wu, D.; Gong, P.; Gao, M. Crystal structure of Bacillus thuringiensis Cry7Ca1 toxin active against Locusta migratoria manilensis. Protein Sci. 2019, 28, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Evdokimov, A.G.; Moshiri, F.; Sturman, E.J.; Rydel, T.J.; Zheng, M.; Seale, J.W.; Franklin, S. Structure of the full-length insecticidal protein Cry1Ac reveals intriguing details of toxin packaging into in vivo formed crystals. Protein Sci. 2014, 23, 1491–1497. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, L.E.; Perez, C.; Segovia, L.; Rodriguez, M.H.; Gill, S.S.; Bravo, A.; Soberon, M. Cry11Aa toxin from Bacillus thuringiensis binds its receptor in Aedes aegypti mosquito larvae through loop alpha-8 of domain II. FEBS Lett. 2005, 579, 3508–3514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, L.; Munoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [Green Version]
- Jurat-Fuentes, J.L.; Crickmore, N. Specificity determinants for Cry insecticidal proteins: Insights from their mode of action. J. Invertebr. Pathol. 2017, 142, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Walters, F.S.; Stacy, C.M.; Lee, M.K.; Palekar, N.; Chen, J.S. An engineered chymotrypsin/cathepsin G site in domain I renders Bacillus thuringiensis Cry3A active against Western corn rootworm larvae. Appl. Environ. Microbiol. 2008, 74, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Du, C.; Martin, P.A.; Nickerson, K.W. Comparison of Disulfide Contents and Solubility at Alkaline pH of Insecticidal and Noninsecticidal Bacillus thuringiensis Protein Crystals. Appl. Environ. Microbiol. 1994, 60, 3847–3853. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.L.; Soccol, V.T.; Soccol, C.R. Bacillus thuringiensis: Mechanism of action, resistance, and new applications: A review. Crit. Rev. Biotechnol. 2016, 36, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Pigott, C.R.; Ellar, D.J. Role of receptors in Bacillus thuringiensis crystal toxin activity. Microbiol. Mol. Biol. Rev. 2007, 71, 255–281. [Google Scholar] [CrossRef] [Green Version]
- Tay, W.T.; Mahon, R.J.; Heckel, D.G.; Walsh, T.K.; Downes, S.; James, W.J.; Lee, S.F.; Reineke, A.; Williams, A.K.; Gordon, K.H. Insect Resistance to Bacillus thuringiensis Toxin Cry2Ab Is Conferred by Mutations in an ABC Transporter Subfamily A Protein. PLoS Genet. 2015, 11, e1005534. [Google Scholar] [CrossRef] [PubMed]
- Vachon, V.; Paradis, M.J.; Marsolais, M.; Schwartz, J.L.; Laprade, R. Ionic permeabilities induced by Bacillus thuringiensis in Sf9 cells. J. Membr. Biol. 1995, 148, 57–63. [Google Scholar] [CrossRef]
- Soberon, M.; Portugal, L.; Garcia-Gomez, B.I.; Sanchez, J.; Onofre, J.; Gomez, I.; Pacheco, S.; Bravo, A. Cell lines as models for the study of Cry toxins from Bacillus thuringiensis. Insect. Biochem. Mol. Biol. 2018, 93, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Guihard, G.; Vachon, V.; Laprade, R.; Schwartz, J.L. Kinetic properties of the channels formed by the bacillus thuringiensis insecticidal crystal protein Cry1C in the plasma membrane of Sf9 cells. J. Membr. Biol. 2000, 175, 115–122. [Google Scholar] [CrossRef]
- Portugal, L.; Munoz-Garay, C.; Martinez de Castro, D.L.; Soberon, M.; Bravo, A. Toxicity of Cry1A toxins from Bacillus thuringiensis to CF1 cells does not involve activation of adenylate cyclase/PKA signaling pathway. Insect. Biochem. Mol. Biol. 2017, 80, 21–31. [Google Scholar] [CrossRef]
- Bravo, A.; Gill, S.S.; Soberon, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, C.; Luthy, P.; Hutter, R.; Pliska, V. Binding of the delta endotoxin from Bacillus thuringiensis to brush-border membrane vesicles of the cabbage butterfly (Pieris brassicae). Eur. J. Biochem. 1988, 173, 85–91. [Google Scholar] [CrossRef]
- Hofmann, C.; Vanderbruggen, H.; Hofte, H.; Van Rie, J.; Jansens, S.; Van Mellaert, H. Specificity of Bacillus thuringiensis delta-endotoxins is correlated with the presence of high-affinity binding sites in the brush border membrane of target insect midguts. Proc. Natl. Acad. Sci. USA 1988, 85, 7844–7848. [Google Scholar] [CrossRef] [Green Version]
- Nishiguchi, S.; Yagi, A.; Sakai, N.; Oda, H. Divergence of structural strategies for homophilic E-cadherin binding among bilaterians. J. Cell Sci. 2016, 129, 3309–3319. [Google Scholar] [CrossRef] [Green Version]
- Edelman, R.; Hardegree, M.C.; Chedid, L. From the National Institute of Allergy and Infectious Diseases. Summary of an International Symposium on potentiation of the immune response to vaccines. J. Infect. Dis. 1980, 141, 103–112. [Google Scholar] [CrossRef]
- Jennings, R.; Simms, J.R.; Heath, A.W. Adjuvants and delivery systems for viral vaccines--mechanisms and potential. Dev. Biol. Stand. 1998, 92, 19–28. [Google Scholar]
- Panisset, M. Gaston Ramon decouvrait les anatoxines. Can. J. Comp. Med. Vet. Sci. 1949, 13, 60–63. [Google Scholar] [PubMed]
- Marrack, P.; McKee, A.S.; Munks, M.W. Towards an understanding of the adjuvant action of aluminium. Nat. Rev. Immunol. 2009, 9, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Pasquale, A.; Preiss, S.; Tavares Da Silva, F.; Garcon, N. Vaccine Adjuvants: From 1920 to 2015 and Beyond. Vaccines 2015, 3, 320–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyle, P.M. Biotechnology approaches to produce potent, self-adjuvanting antigen-adjuvant fusion protein subunit vaccines. Biotechnol. Adv. 2017, 35, 375–389. [Google Scholar] [CrossRef]
- Cardenas-Vargas, A.; Elizondo-Quiroga, D.; Gutierrez-Ortega, A.; Charles-Nino, C.; Pedroza-Roldan, C. Evaluation of the Immunogenicity of a Potyvirus-Like Particle as an Adjuvant of a Synthetic Peptide. Viral. Immunol. 2016, 29, 557–564. [Google Scholar] [CrossRef]
- Petrovsky, N.; Aguilar, J.C. Vaccine adjuvants: Current state and future trends. Immunol. Cell Biol. 2004, 82, 488–496. [Google Scholar] [CrossRef]
- Clements, C.J.; Griffiths, E. The global impact of vaccines containing aluminium adjuvants. Vaccine 2002, 20 (Suppl. 3), S24–S33. [Google Scholar] [CrossRef]
- Banzhoff, A.; Gasparini, R.; Laghi-Pasini, F.; Staniscia, T.; Durando, P.; Montomoli, E.; Capecchi, P.L.; di Giovanni, P.; Sticchi, L.; Gentile, C.; et al. MF59-adjuvanted H5N1 vaccine induces immunologic memory and heterotypic antibody responses in non-elderly and elderly adults. PLoS ONE 2009, 4, e4384. [Google Scholar] [CrossRef]
- Barker, C.I.; Snape, M.D. Pandemic influenza A H1N1 vaccines and narcolepsy: Vaccine safety surveillance in action. Lancet Infect. Dis. 2014, 14, 227–238. [Google Scholar] [CrossRef]
- Morel, S.; Didierlaurent, A.; Bourguignon, P.; Delhaye, S.; Baras, B.; Jacob, V.; Planty, C.; Elouahabi, A.; Harvengt, P.; Carlsen, H.; et al. Adjuvant System AS03 containing alpha-tocopherol modulates innate immune response and leads to improved adaptive immunity. Vaccine 2011, 29, 2461–2473. [Google Scholar] [CrossRef] [PubMed]
- Moyo-Gwete, T.; Madzivhandila, M.; Makhado, Z.; Ayres, F.; Mhlanga, D.; Oosthuysen, B.; Lambson, B.E.; Kgagudi, P.; Tegally, H.; Iranzadeh, A.; et al. SARS-CoV-2 501Y.V2 (B.1.351) elicits cross-reactive neutralizing antibodies. bioRxiv 2021. [Google Scholar] [CrossRef]
- Lawson, L.B.; Norton, E.B.; Clements, J.D. Defending the mucosa: Adjuvant and carrier formulations for mucosal immunity. Curr. Opin. Immunol. 2011, 23, 414–420. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, N.; Teixeira, J.; Roteli-Martins, C.M.; Naud, P.; De Borba, P.; Zahaf, T.; Sanchez, N.; Schuind, A. Sustained efficacy and immunogenicity of the HPV-16/18 AS04-adjuvanted vaccine up to 7.3 years in young adult women. Vaccine 2010, 28, 6247–6255. [Google Scholar] [CrossRef] [PubMed]
- Garcon, N.; Morel, S.; Didierlaurent, A.; Descamps, D.; Wettendorff, M.; Van Mechelen, M. Development of an AS04-adjuvanted HPV vaccine with the adjuvant system approach. BioDrugs 2011, 25, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Kirschning, C.J.; Wesche, H.; Merrill Ayres, T.; Rothe, M. Human toll-like receptor 2 confers responsiveness to bacterial lipopolysaccharide. J. Exp. Med. 1998, 188, 2091–2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dauphinee, S.M.; Karsan, A. Lipopolysaccharide signaling in endothelial cells. Lab. Invest. 2006, 86, 9–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hagan, D.T.; Valiante, N.M. Recent advances in the discovery and delivery of vaccine adjuvants. Nat. Rev. Drug Discov. 2003, 2, 727–735. [Google Scholar] [CrossRef]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Spangler, B.D. Structure and function of cholera toxin and the related Escherichia coli heat-labile enterotoxin. Microbiol. Rev. 1992, 56, 622–647. [Google Scholar] [CrossRef]
- Clements, J.D.; Hartzog, N.M.; Lyon, F.L. Adjuvant activity of Escherichia coli heat-labile enterotoxin and effect on the induction of oral tolerance in mice to unrelated protein antigens. Vaccine 1988, 6, 269–277. [Google Scholar] [CrossRef]
- Lycke, N.; Holmgren, J. Strong adjuvant properties of cholera toxin on gut mucosal immune responses to orally presented antigens. Immunology 1986, 59, 301–308. [Google Scholar]
- Ge, A.Z.; Pfister, R.M.; Dean, D.H. Hyperexpression of a Bacillus thuringiensis delta-endotoxin-encoding gene in Escherichia coli: Properties of the product. Gene 1990, 93, 49–54. [Google Scholar] [CrossRef]
- Rodriguez-Orozco, A.R.; Ayala-Mata, F.; Tinoco-Martinez, R.; Cabrera-Navarro, L. The new mucosal adjuvant Cry1Ac increase the capacity of mononuclear cells to stop bacterial growth. Rev. Invest. Clin. 2007, 59, 161–163. [Google Scholar]
- Vazquez, R.I.; Moreno-Fierros, L.; Neri-Bazan, L.; De La Riva, G.A.; Lopez-Revilla, R. Bacillus thuringiensis Cry1Ac protoxin is a potent systemic and mucosal adjuvant. Scand. J. Immunol. 1999, 49, 578–584. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Fierros, L.; Garcia, N.; Gutierrez, R.; Lopez-Revilla, R.; Vazquez-Padron, R.I. Intranasal, rectal and intraperitoneal immunization with protoxin Cry1Ac from Bacillus thuringiensis induces compartmentalized serum, intestinal, vaginal and pulmonary immune responses in BALB/c mice. Microbes. Infect. 2000, 2, 885–890. [Google Scholar] [CrossRef]
- Moreno-Fierros, L.; Ruiz-Medina, E.J.; Esquivel, R.; Lopez-Revilla, R.; Pina-Cruz, S. Intranasal Cry1Ac protoxin is an effective mucosal and systemic carrier and adjuvant of Streptococcus pneumoniae polysaccharides in mice. Scand. J. Immunol. 2003, 57, 45–55. [Google Scholar] [CrossRef]
- Parenti, M.D.; Santoro, A.; Del Rio, A.; Franceschi, C. Literature review in support of adjuvanticity/immunogenicity assessment of proteins. EFSA Supporting Publ. 2019, 16, 1551E. [Google Scholar] [CrossRef]
- Lycke, N. Recent progress in mucosal vaccine development: Potential and limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef]
- Bomsel, M.; Tudor, D.; Drillet, A.S.; Alfsen, A.; Ganor, Y.; Roger, M.G.; Mouz, N.; Amacker, M.; Chalifour, A.; Diomede, L.; et al. Immunization with HIV-1 gp41 subunit virosomes induces mucosal antibodies protecting nonhuman primates against vaginal SHIV challenges. Immunity 2011, 34, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, V.; Drumare, M.F.; Lereclus, D.; Gohar, M.; Lamourette, P.; Nevers, M.C.; Vaisanen-Tunkelrott, M.L.; Bernard, H.; Guillon, B.; Creminon, C.; et al. In vitro digestion of Cry1Ab proteins and analysis of the impact on their immunoreactivity. J Agric. Food Chem. 2010, 58, 3222–3231. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Hernandez, S.; Rodriguez-Monroy, M.A.; Lopez-Revilla, R.; Resendiz-Albor, A.A.; Moreno-Fierros, L. Intranasal coadministration of the Cry1Ac protoxin with amoebal lysates increases protection against Naegleria fowleri meningoencephalitis. Infect. Immun. 2004, 72, 4368–4375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Martinez, M.; Rubio-Infante, N.; Garcia-Hernandez, A.L.; Nava-Acosta, R.; Ilhuicatzi-Alvarado, D.; Moreno-Fierros, L. Cry1Ac toxin induces macrophage activation via ERK1/2, JNK and p38 mitogen-activated protein kinases. Int. J. Biochem. Cell Biol. 2016, 78, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.S.; Barnett, B.; Doerrer, N.G.; Glenn, K.; Herman, R.A.; Herouet-Guicheney, C.; Hunst, P.; Kough, J.; Ladics, G.S.; McClain, S.; et al. Assessment of potential adjuvanticity of Cry proteins. Regul. Toxicol. Pharmacol. 2016, 79, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Moreno, S.; Garcia-Hernandez, A.L.; Moreno-Fierros, L. Coadministration of protoxin Cry1Ac from Bacillus thuringiensis with metacestode extract confers protective immunity to murine cysticercosis. Parasite Immunol. 2014, 36, 266–270. [Google Scholar] [CrossRef]
- Gonzalez-Gonzalez, E.; Garcia-Hernandez, A.L.; Flores-Mejia, R.; Lopez-Santiago, R.; Moreno-Fierros, L. The protoxin Cry1Ac of Bacillus thuringiensis improves the protection conferred by intranasal immunization with Brucella abortus RB51 in a mouse model. Vet. Microbiol. 2015, 175, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Legorreta-Herrera, M.; Meza, R.O.; Moreno-Fierros, L. Pretreatment with Cry1Ac protoxin modulates the immune response, and increases the survival of Plasmodium-infected CBA/Ca mice. J. Biomed. Biotechnol. 2010, 2010, 198921. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Padron, R.I.; Gonzales-Cabrera, J.; Garcia-Tovar, C.; Neri-Bazan, L.; Lopez-Revilla, R.; Hernandez, M.; Moreno-Fierro, L.; de la Riva, G.A. Cry1Ac protoxin from Bacillus thuringiensis sp. kurstaki HD73 binds to surface proteins in the mouse small intestine. Biochem. Biophys. Res. Commun. 2000, 271, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, G.G.; Dean, D.H.; Moreno-Fierros, L. Structural implication of the induced immune response by Bacillus thuringiensis Cry proteins: Role of the N-terminal region. Mol. Immunol. 2004, 41, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Adel-Patient, K.; Guimaraes, V.D.; Paris, A.; Drumare, M.F.; Ah-Leung, S.; Lamourette, P.; Nevers, M.C.; Canlet, C.; Molina, J.; Bernard, H.; et al. Immunological and metabolomic impacts of administration of Cry1Ab protein and MON 810 maize in mouse. PLoS ONE 2011, 6, e16346. [Google Scholar] [CrossRef]
- Wilson-Welder, J.H.; Torres, M.P.; Kipper, M.J.; Mallapragada, S.K.; Wannemuehler, M.J.; Narasimhan, B. Vaccine adjuvants: Current challenges and future approaches. J. Pharm. Sci. 2009, 98, 1278–1316. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Infante, N.; Moreno-Fierros, L. An overview of the safety and biological effects of Bacillus thuringiensis Cry toxins in mammals. J. Appl. Toxicol. 2016, 36, 630–648. [Google Scholar] [CrossRef] [PubMed]
| 1 |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cry Protein | Pests Controlled |
|---|---|
| Cry1A, Cry2A, Cry3A, Cry14A | Coleoptera |
| Cry1Ac, Cry2A | Lepidoptera |
| Cry1A, Cry2A, Cry4A, Cry10A | Diptera |
| Cry2A, Cry3A, Cry11A | Hemiptera |
| Cry3A, Cry5A, Cry22A | Hymenoptera |
| Human Cadherin () 1 | Insect Cadherin 1 | Identity % |
|---|---|---|
| Type I (EAW83244.1) | Drosophila melanogaster (ACD79974.1) Manduca sexta (AAM21151.1) Anopheles gambiae (AGN95449.1) Pieris rapae (XP_022120264.1) Bombyx mori (XP_ 012545103.1) Tribolium castaneum (EEZ99177.2) Aedes aegypti (XP_021693027.1) | 14.94 14.30 13.51 18.71 20.45 20.30 19.63 |
| E-cadherin (CAA78353.1) | Drosophila melanogaster (ACD79974.1) Manduca sexta (AAM21151.1) Anopheles gambiae (AGN95449.1) Pieris rapae (XP_022120264.1) Bombyx mori (XP_ 012545103.1) Tribolium castaneum (EEZ99177.2) Aedes aegypti (XP_021693027.1) | 14.42 14.03 13.29 18.43 20.51 20.14 19.74 |
| Adjuvant Composition | Immune Mechanism | Use in Vaccines | References |
|---|---|---|---|
| Aluminum salts (alum salts, aluminum hydroxide, aluminum phosphate, and aluminum sulfate phosphate) | Activation of the Nalp3/NLRP3 complex, leading to a considerable increase in IL-1β and IL-8. An increase in the chemokines CCL2, CCL3, and CCL4, which activate macrophages, has also been observed. | Diphtheria, pertussis, tetanus, hepatitis A and B viruses, meningococci, and human papillomavirus (HPV) | [69,72,73,74] |
| MF59 (combination of squalene, Polysorbate 80, and Span 85) | Directly induces the arrival of inflammatory cells, such as macrophages, and promotes the secretion of chemokines such as CCL4, CCL5, and CCL12. It can also activate Th1-type cytokines. | Simplex herpes virus, human immunodeficiency, and seasonal influenza | [72,75] |
| ASO3 (combination of squalene, vitamin E, and polysorbate) | Induces the arrival of macrophages, which secrete cytokines such as IL-6. | H5N1 and H1N1 influenza vaccines COVID-19 1 | [76,77] [78] |
| ASO4 (combination of monophosphoryl lipid A and aluminum salts) | TLR4-expressing cells in the muscle, such as resident or recruited dendritic cells or monocytes, are activated and induce the rapid recruitment and activation of monocytes and dendritic cells. | Hepatitis B virus (HBV) and HPV | [79,80,81] |
| Lipopolysaccharide (LPS) | Interacts directly on the TLR4 receptor, which can induce the secretion of cytokines TNF-α and IL-6 as well as various chemokines. | HPV and HBV | [82,83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez-Vazquez, M.C.; Vela-Sanchez, R.A.; Rojas-Ruiz, N.E.; Carabarin-Lima, A. Importance of Cry Proteins in Biotechnology: Initially a Bioinsecticide, Now a Vaccine Adjuvant. Life 2021, 11, 999. https://doi.org/10.3390/life11100999
Gonzalez-Vazquez MC, Vela-Sanchez RA, Rojas-Ruiz NE, Carabarin-Lima A. Importance of Cry Proteins in Biotechnology: Initially a Bioinsecticide, Now a Vaccine Adjuvant. Life. 2021; 11(10):999. https://doi.org/10.3390/life11100999
Chicago/Turabian StyleGonzalez-Vazquez, Maria Cristina, Ruth Abril Vela-Sanchez, Norma Elena Rojas-Ruiz, and Alejandro Carabarin-Lima. 2021. "Importance of Cry Proteins in Biotechnology: Initially a Bioinsecticide, Now a Vaccine Adjuvant" Life 11, no. 10: 999. https://doi.org/10.3390/life11100999
APA StyleGonzalez-Vazquez, M. C., Vela-Sanchez, R. A., Rojas-Ruiz, N. E., & Carabarin-Lima, A. (2021). Importance of Cry Proteins in Biotechnology: Initially a Bioinsecticide, Now a Vaccine Adjuvant. Life, 11(10), 999. https://doi.org/10.3390/life11100999